
Abstract
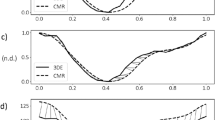
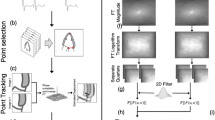
Technological advances in image quality and post-processing have led to the better clinical adoption of 3D echocardiography to quantify cardiac function. However, the right ventricle (RV) raises specific challenges due to its specific half-moon shape, which led to a lack of consensus regarding the estimation of RV motion and deformation locally. In this paper, we detail three ways to estimate local anatomically-relevant directions at each point of the RV surface, in 3D, and the resulting Green-Lagrange strain projected along these directions. Using a database of RV surface meshes extracted from 3D echocardiographic sequences from 100 control subjects, we quantified differences between these strategies in terms of local anatomical directions and local strain, both at the individual and population levels. For the latter, we used a specific dimensionality reduction technique to align the latent spaces encoding the strain patterns obtained from different computations of the anatomical directions. Differences were subtle but visible at specific regions of the RV and partially interpretable, although their impact on the population latent representation was low, which sets a preliminary quantitative basis to discuss these computation standards.
Access this chapter
Tax calculation will be finalised at checkout
Purchases are for personal use only
Similar content being viewed by others

References
Sanz, J., Sánchez-Quintana, D., Bossone, E., Bogaard, H.J., Naeije, R.: Anatomy, function, and dysfunction of the right ventricle: JACC state-of-the-art review. J. Am. Coll. Cardiol. 73, 1463–82 (2019)
Addetia, K., Muraru, D., Badano, L.P., Lang, R.M.: New directions in right ventricular assessment using 3-dimensional echocardiography. JAMA Cardiol. 4, 936–44 (2019)
Duchateau, N., Moceri, P., Sermesant, M.: Direction-dependent decomposition of 3D right ventricular motion: beware of approximations. J. Am. Soc. Echocardiogr. 34, 201–3 (2021)
D’hooge, J., et al.: Regional strain and strain rate measurements by cardiac ultrasound: principles, implementation and limitations. Eur. J. Echocardiogr. 1, 154–70 (2000)
Lakatos, B.K., Nabeshima, Y., Tokodi, M., Nagata, Y., Tösér, Z., Otani, K., et al.: Importance of nonlongitudinal motion components in right ventricular function: three-dimensional echocardiographic study in healthy volunteers. J. Am. Soc. Echocardiogr. 33, 995-1005.e1 (2020)
Tokodi, M., Staub, L., Budai, A., Lakatos, B.K., Csákvári, F.I., Suhai, M., et al.: Partitioning the right ventricle into 15 segments and decomposing its motion using 3D echocardiography-based models: the updated ReVISION method. Front. Cardiovasc. Med. 8, 622118 (2021)
Moceri, P., Duchateau, N., Gillon, S., Jaunay, L., Baudouy, D., Squara, F., et al.: Three-dimensional right ventricular shape and strain in congenital heart disease patients with right ventricular chronic volume loading. Eur. Heart J. Cardiovasc. Imaging 22, 1174–81 (2021)
Satriano, A., Pournazari, P., Hirani, N., Helmersen, D., Thakrar, M., Weatherald, J., et al.: Characterization of right ventricular deformation in pulmonary arterial hypertension using three-dimensional principal strain analysis. J. Am. Soc. Echocardiogr. 32, 385–93 (2019)
Mitchell, J., Mount, D., Papadimitriou, C.: The discrete geodesic problem. SIAM J. Comput. 16, 647–68 (1987)
Di Folco, M., Moceri, P., Clarysse, P., Duchateau, N.: Characterizing interactions between cardiac shape and deformation by non-linear manifold learning. Med. Image Anal. 75, 102278 (2022)
Belkin, M., Niyogi, P.: Laplacian eigenmaps for dimensionality reduction and data representation. Neural Comput. 15, 1373–96 (2003)
Acknowledgements
The authors acknowledge the support from the French ANR (LABEX PRIMES of Univ. Lyon [ANR-11-LABX-0063], the JCJC project “MIC-MAC” [ANR-19-CE45-0005]), and the European Union - NextGenerationEU, Ministry of Universities and Recovery, Transformation and Resilience Plan, through a call from Pompeu Fabra University (Barcelona). They are also grateful to P. Moceri (CHU Nice, France) for providing the imaging data related to the studied population, and to M. Sermesant (INRIA Epione, Sophia Antipolis, France) for initial discussions on this topic.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2023 The Author(s), under exclusive license to Springer Nature Switzerland AG
About this paper

Cite this paper
Di Folco, M., Dargent, T., Bernardino, G., Clarysse, P., Duchateau, N. (2023). Which Anatomical Directions to Quantify Local Right Ventricular Strain in 3D Echocardiography?. In: Bernard, O., Clarysse, P., Duchateau, N., Ohayon, J., Viallon, M. (eds) Functional Imaging and Modeling of the Heart. FIMH 2023. Lecture Notes in Computer Science, vol 13958. Springer, Cham. https://doi.org/10.1007/978-3-031-35302-4_62
Download citation
DOI: https://doi.org/10.1007/978-3-031-35302-4_62
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-031-35301-7
Online ISBN: 978-3-031-35302-4
eBook Packages: Computer ScienceComputer Science (R0)