
Abstract
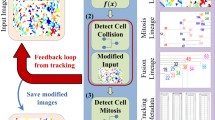
We present a novel framework for high-throughput cell lineage analysis in time-lapse microscopy images. Our algorithm ties together two fundamental aspects of cell lineage construction, namely cell segmentation and tracking, via a Bayesian inference of dynamic models. The proposed contribution exploits the Kalman inference problem by estimating the time-wise cell shape uncertainty in addition to cell trajectory. These inferred cell properties are combined with the observed image measurements within a fast marching (FM) algorithm, to achieve posterior probabilities for cell segmentation and association. Highly accurate results on two different cell-tracking datasets are presented.
Chapter PDF
Similar content being viewed by others

Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
References
Amat, F., Lemon, W., Mossing, D.P., McDole, K., Wan, Y., Branson, K., Myers, E.W., Keller, P.J.: Fast, accurate reconstruction of cell lineages from large-scale fluorescence microscopy data. Nature Methods (2014)
Carpenter, A.E., Jones, T.R., Lamprecht, M.R., Clarke, C., Kang, I.H., Friman, O., Guertin, D.A., Chang, J.H., Lindquist, R.A., Moffat, J., et al.: Cellprofiler: image analysis software for identifying and quantifying cell phenotypes. Genome Biology 7(10), R100 (2006)
Cohen, A.A., Geva-Zatorsky, N., Eden, E., Frenkel-Morgenstern, M., Issaeva, I., Sigal, A., Milo, R., Cohen-Saidon, C., Liron, Y., Kam, Z., et al.: Dynamic proteomics of individual cancer cells in response to a drug. Science 322(5907), 1511–1516 (2008)
Dzyubachyk, O., van Cappellen, W.A., Essers, J., Niessen, W.J., Meijering, E.: Advanced level-set-based cell tracking in time-lapse fluorescence microscopy. IEEE Trans. on Medical Imaging 29(3), 852–867 (2010)
Gilad, T., Bray, M., Carpenter, A., Riklin-Raviv, T.: Symmetry-based mitosis detection in time-lapse microscopy. In: IEEE International Symposium on Biomedical Imaging: From Nano to Macro (2015)
Hassouna, M.S., Farag, A.A.: Multistencils fast marching methods: A highly accurate solution to the eikonal equation on cartesian domains. IEEE Trans. on Pattern Analysis and Machine Intelligence 29(9), 1563–1574 (2007)
Jaqaman, K., Loerke, D., Mettlen, M., Kuwata, H., Grinstein, S., Schmid, S.L., Danuser, G.: Robust single-particle tracking in live-cell time-lapse sequences. Nature Methods 5(8), 695–702 (2008)
Kachouie, N.N., Fieguth, P.W.: Extended-hungarian-jpda: Exact single-frame stem cell tracking. IEEE Trans. Biomed. Eng. 54(11), 2011–2019 (2007)
Kalman, R.E.: A new approach to linear filtering and prediction problems. J. of Fluids Engineering 82(1), 35–45 (1960)
Kanade, T., Yin, Z., Bise, R., Huh, S., Eom, S., Sandbothe, M.F., Chen, M.: Cell image analysis: Algorithms, system and applications. In: 2011 IEEE Workshop on Applications of Computer Vision (WACV), pp. 374–381. IEEE (2011)
Maška, M., Ulman, V., Svoboda, D., Matula, P., Matula, P., Ederra, C., Urbiola, A., España, T., Venkatesan, S., Balak, D.M., et al.: A benchmark for comparison of cell tracking algorithms. Bioinformatics 30(11), 1609–1617 (2014)
Meijering, E., Dzyubachyk, O., Smal, I., et al.: Methods for cell and particle tracking. Methods Enzymol. 504(9), 183–200 (2012)
Padfield, D., Rittscher, J., Roysam, B.: Spatio-temporal cell segmentation and tracking for automated screening. In: IEEE International Symposium on Biomedical Imaging: From Nano to Macro, pp. 376–379. IEEE (2008)
Padmanabhan, K., Eddy, W.F., Crowley, J.C.: A novel algorithm for optimal image thresholding of biological data. J. of Neuroscience Methods 193(2), 380–384 (2010)
Pohl, K.M., Fisher, J., Shenton, M.E., McCarley, R.W., Grimson, W.E.L., Kikinis, R., Wells, W.M.: Logarithm odds maps for shape representation. In: Larsen, R., Nielsen, M., Sporring, J. (eds.) MICCAI 2006. LNCS, vol. 4191, pp. 955–963. Springer, Heidelberg (2006)
Rapoport, D.H., Becker, T., Mamlouk, A.M., Schicktanz, S., Kruse, C.: A novel validation algorithm allows for automated cell tracking and the extraction of biologically meaningful parameters. PloS One 6(11), e27315 (2011)
Reid, D.B.: An algorithm for tracking multiple targets. IEEE Transactions on Automatic Control 24(6), 843–854 (1979)
Riklin-Raviv, T., Van Leemput, K., Menze, B.H., Wells, W.M., Golland, P.: Segmentation of image ensembles via latent atlases. Medical Image Analysis 14(5), 654–665 (2010)
Schiegg, M., Hanslovsky, P., Haubold, C., Koethe, U., Hufnagel, L., Hamprecht, F.A.: Graphical model for joint segmentation and tracking of multiple dividing cells. Bioinformatics, page btu764 (2014)
Su, H., Yin, Z., Huh, S., Kanade, T.: Cell segmentation in phase contrast microscopy images via semi-supervised classification over optics-related features. Medical Image Analysis 17(7), 746–765 (2013)
Yang, X., Li, H., Zhou, X.: Nuclei segmentation using marker-controlled watershed, tracking using mean-shift, and kalman filter in time-lapse microscopy. IEEE Trans. Circuits Syst. I, Reg. Papers 53(11), 2405–2414 (2006)
Author information
Authors and Affiliations
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2015 Springer International Publishing Switzerland
About this paper
Cite this paper
Arbelle, A., Drayman, N., Bray, M., Alon, U., Carpenter, A., Raviv, T.R. (2015). Analysis of High-throughput Microscopy Videos: Catching Up with Cell Dynamics. In: Navab, N., Hornegger, J., Wells, W., Frangi, A. (eds) Medical Image Computing and Computer-Assisted Intervention – MICCAI 2015. MICCAI 2015. Lecture Notes in Computer Science(), vol 9351. Springer, Cham. https://doi.org/10.1007/978-3-319-24574-4_26
Download citation
DOI: https://doi.org/10.1007/978-3-319-24574-4_26
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-24573-7
Online ISBN: 978-3-319-24574-4
eBook Packages: Computer ScienceComputer Science (R0)